
Professor
Ph.D., Georgia Institute of Technology, 1986
Biochemistry
Tel: 316.978.7386
Fax: 316.978.3431
Email: Kandatege.Wimalasena@wichita.edu
Dr. Wimalasena's Research Group
Research Interests
Molecular Causes of the Specific Degeneration of the Dopaminergic System in PD
Although the exact cause(s) of dopaminergic cell death in Parkinson's disease (PD)
is not understood, the commonly accepted view is that the exposure to environmental
toxins may be a major cause of PD, especially in individuals with cumulative defects
in pathways associated with handling of oxidative stress, mitochondrial function,
calcium homeostasis, and the ubiquitin proteasome system. The discovery that 1-methyl-4-phenylpyridinium
(MPP+) selectively destroys dopaminergic neurons and causes PD symptoms in humans and other
mammals has strengthened the environmental hypothesis of PD. Therefore, MPP+ has been extensively used to model the environmental causes of PD. The current model
for the selective dopaminergic toxicity of MPP+ is centered on the specific cellular uptake into dopaminergic cells through the plasma
membrane dopamine transporter (DAT) , complex I inhibition leading to ATP depletion,
increased reactive oxygen species (ROS) production, and finally apoptotic cell death.
However, we have shown that MPP+ is taken up not only into dopaminergic cells, but also into various other cells,
including liver cells through various other transporters with high efficiency. Thus,
the selective dopaminergic toxicity of MPP+ could not be due to the specific uptake into dopaminergic cells through DAT. The
focus of current research in my lab is to determine the molecular causes of specific
degeneration of the dopaminergic system in PD. Our specific research interested are
as follows.
(A) The mechanism of the selective vulnerability of dopaminergic neurons towards Parkinsonian
mitochondrial toxins similar to MPP+
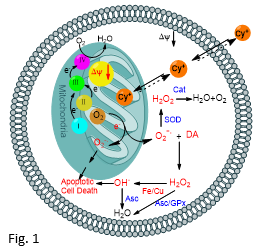
Based on our recent in vitro studies using a series of novel lipophilic mitochondrial toxins, we proposed that the selective high vulnerability of dopaminergic neurons towards mitochondrial toxins similar to MPP+ must be due to their intrinsic characteristics, including, presence of high levels of oxidatively labile dopamine and relatively low levels of antioxidant enzymes, greater demand for energy, etc. (Fig. 1).1-3 Current research is focused on the extension of these in vitro findings to appropriate in vivo systems.
(B) Role of Ca2+ on Mitochondrial uptake and toxicity of MPP+:
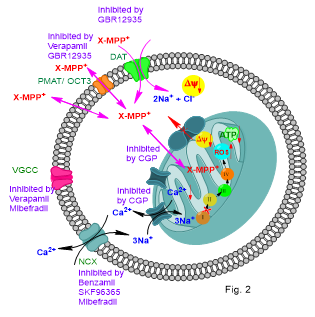
The objective of this study was to characterize structural and functional MPP+ mimics suitable to study the cellular distribution and mitochondrial uptake of MPP+ in live cells, and use them to characterize the cellular mechanisms involved in these processes to advance the understanding of the mechanism of the selective dopaminergic cell toxicity of MPP+. Using these novel probes, we have recently shown that the extra-mitochondrial Ca2+ and the specific mitochondrial sodium calcium-exchanger (mNCX) inhibitor, CGP37157, inhibits the mitochondrial uptake and that CGP37157 protects dopaminergic MN9D cells from MPP+ toxicity. Therefore, we propose that mNCX could be a potential target for the development of pharmaceuticals for the protection of the CNS dopaminergic neurons from environmental toxins similar to MPP+ (Fig. 2).4 We are currently exploring these possibilities.
(C) Does intracellular acidosis mediated TH activation play a role in selective dopaminergic
cell death in PD?
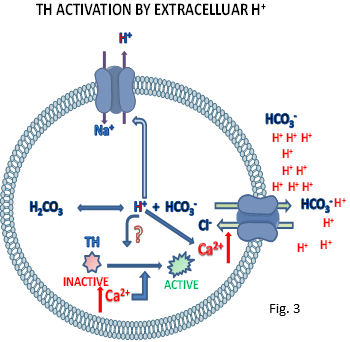
Catecholamines cause high oxidative stress, especially when their tightly regulated homeostasis is perturbed. Thus, unwanted activation of the tightly regulated tyrosine hydroxylase (TH), the rate limiting enzyme of the catecholamine biosynthetic pathway, leads to the over production of catecholamines which could cause high oxidative stress and cell death. We have recently discovered that intracellular acidosis results in an unprecedented increase of TH activity in dopaminergic MN9D and adrenergic PC12 cells [unpublished results; (Fig. 3)]. Based on these findings, together with the finding that acidosis is commonly associated with many types of CNS neurodegenerative conditions, we propose that intracellular acidosis mediated TH activation may contribute to the high vulnerability of catecholamergic neurons in these diseases. In current studies, we plan to experimentally test this proposal in vitro using rat embryonic mesencephalic and human stem cell derived dopaminergic neuronal cultures.
References
- Kadigamuwa, C. C., Mapa, S. T., Wimalasena, K. “Lipophilic Cationic Cyanines Are Potent Complex I Inhibitors and Specific in Vitro Dopaminergic Toxins with Mechanistic Similarities to Both Rotenone and MPP+” Chem. Res. Toxicol. (ACS), 2016, 28, 1468-1479.
- Wimalasena K. “The inherent high vulnerability of dopaminergic neurons towards mitochondrial toxins may contribute to the etiology of Parkinson’s disease (PD)” Neural Regeneration Research 2016, 11, 246-247 Invited Perspective.
- Kadigamuwa, C. C., Le, V. Q., Wimalasena, K. “ 2,2- and 4, 4-Cyanines are transporter independent In vivo dopaminergic toxins with the specificity and mechanism of toxicity similar to MPP+” J. Neurochemistry, 2015, 135, 755-767.
- Mapa, S. T., Le, V. Q., Wimalasena, K. “Characteristics of the mitochondrial uptake of Parkinsonian toxin, MPP+ as probed by the fluorescent mimic, 4'I-MPP+ in live cells” Chem. Res. Toxicol. (ACS), 2017 (submitted).